5.
Haplogrupa R1a i jej podgrupy R1a1a i R1a1a7.
Jak, gdzie i kiedy wydzieliły sie haplogrupy Słowian
i ich najbliższych krewnych: Bałtów i indo-irańskich Ariów?
Z genezą Słowian jako etnosu (gr. "etnos" - plemię,
ród) sprawa jest niejasna, podobnie jak z innymi starożytnymi ludami. Na
podstawie opisów, jakie zostawili nam starożytni pisarze, ale dopiero
począwszy od Herodota (V w. przed Chr.), Słowianie odznaczali
się trzema głównymi cechami: "Po pierwsze: lokalizacja gdzieś
w dorzeczu Dniepru, Dniestru, po Karpaty lub ewentualnie ujście Dunaju. Po
drugie: usytuowanie na terenach o dobrze rozwiniętej sieci wód
śródlądowych, gęstych lasach, bagnach, na obszarach wybitnie
nizinnych. Po trzecie: muszą być w sensowny sposób związane z
młodszymi kulturami archeologicznymi z okresu wczesnego
średniowiecza, dla których stanowiły podstawę uformowania sie i
dalszego rozwoju" (A. Błażejewski). Na tej podstawie
Słowianom można by przypisać archeologiczne kultury
wysocką i wołyńską nad górnym Bugiem i Dniestrem
(może to dzieła Herodotowych Neurów i Budynów z około V w. przed
Chr.), a już na pewno zarubiniecką i czerniachowską (od III w.
przed Chr.) oraz kijowską (od III w. po Chr.), na Ukrainie
(Niestety, złą
przysługę prawdzie naukowej wyrządzili ostatnio skrajni
allochtoniści, zwani kossinowcami-dnieprowcami, którzy - na podstawie
zacieśnionej definicji archeologicznej kultury słowiańskiej -
początek Słowian na ziemiach datują jedynie na VI w., ich
praojczyznę lokują jedynie nad środkowym i górnym Dnieprem, a
ich genezę ustalają tam na czas niewiele wcześniejszy. Za nimi
poszli niektórzy polscy genetycy z kręgu medycyny sądowej, np. K.
Rębała i zesp. 2007, którzy badając pochodzenie dzisiejszych
Polaków, nie rozróżniają ich chromosomowych haplogrup SNP, a
biorąc pod uwagę tylko haplotypy STR, nieraz bardzo podobne mimo
odległych pochodzeniem haplogrup SNP, oceniają ich podobieństwo
czy różnorodność względem haplotypów innych populacji i na
tej podstawie orzekają o dacie ich genezy w... V czy VI w. i miejscu
ich oddzielenia się od innych ludów - nad Dnieprem! Podobna metoda, oparta
na mieszaniu haplotypów rozmaitych haplogrup i korzystaniu
także z materiału autosomalnego, zawiodła już
wcześniej R. Płoskiego i zesp. 2002 do stwierdzenia, wbrew naukowej
genetyce, o wielkiej odrębności Polaków od innych ludów
słowiańskich; że pod "względem genetycznym
jesteśmy prawie tak samo oddaleni od Moskwy, jak i od Berlina").
Rzecz w
tym jednak, że rzetelna genetyczna genealogia Y-DNA i językoznawstwo
historyczne nie pozwalają na takie, jak wyżej, czasowe, terytorialne
i kulturowe zacieśnienie definicji Słowian starożytnych. Poucza
o tym analiza czasu ich powstania oraz geograficznego rozprzestrzenienia
się populacji Słowian, wyróżniającej się własnym
językiem i genową haplogrupą R1a1a (znana do niedawna jako
R1a1). W listopadzie 2009 z tej haplogrupy
wydzielono ważną podgrupę polską, R1a1a7.
Otóż w populacji hg R1
(M173), oprócz wyżej omówionej "celtyckiej" M343 (haplogrupa
R1b), dokonała się też mutacja M420, dająca
początek haplogrupie R1a, następnie mutacja SRY1532.2,
określana jako haplogrupaR1a1. Te haplogrupy
nie zostawiły po sobie liczniejszych populacji. Natomiast po kolejnych
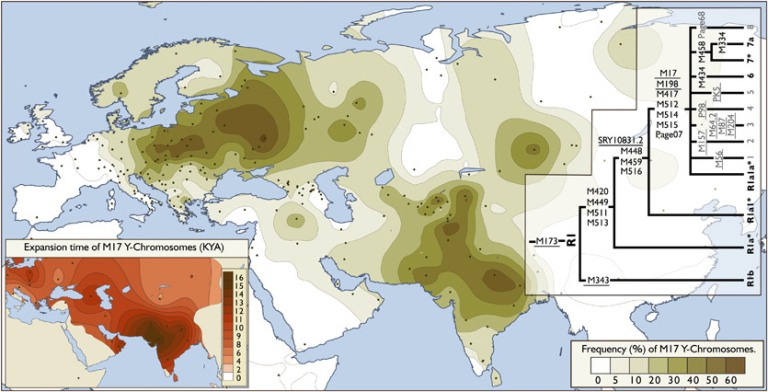
mutacjach (M17 i M198) - powstała wielka eurazyjska, zwłaszcza
indo-słowianska haplogrupa R1a1a, a
wkrótce w jej części europejskiej doszło do mutacji M458,
tworzącej ściśle środkowoeuropejską,
zachodniosłowiańską, głównie polską, haplogrupę R1a1a7.
DRZEWO R, R1 i R1a - według ISOGG
|
R M207/UTY2,
P224, P227, P229, P232, P280, P285, S4, S8, S9, V45 • R* - po kilka procent w niektórych plemionach
Północnego Pakistanu • R1 M173/P241,
M306/S1,P225, P231, P233, P234, P236, P238, P242,
P245, P286, P294 • • R1* - nieliczni w Turcji, Pn.-Wsch. Iranie, Pakistanie, Indiach i w
rejonie Ałtaju. • • R1a L62/M513,
L63/M511, L145/M449, L146/M420 • •
• R1a* - nieliczni na Bliskim Wschodzie, Kaukazie, w Iranie i
Arabii. • •
• R1a1 L120/M516,
L122/M448, M459, SRY1532.2/SRY10831.2 • •
• • R1a1* - nieliczni od Północnej i Pd.-Wsch. Europy do Indii. • •
• • R1a1a M17, M198,
M417, M512, M514, M515, Page07 • •
• •
• R1a1a* - w Azji od około 20000 lat; w Europie od około
12000 lat. • •
• •
• R1a1a1 M56 - pojedyncze próbki • •
• •
• R1a1a2 M157 - gałąź
stara europejska, od ok. 12000 lat • •
• •
• R1a1a3 M64.2,
M87, M204 - gałąź stara
europejska, jak wyżej • •
• •
• R1a1a4 P98 - jedna próbka • •
• •
• R1a1a5 PK5 - jedna próbka w Pakistanie • •
• • • R1a1a6 M434
- nieliczni na Bliskim
Wschodzie i w Pakistanie • •
• •
• R1a1a7 M458 • •
• •
• • R1a1a7* - gałąź N - środkowoeuropejska
(nie-tylko-polska) • •
• •
• • R1a1a7a M334
- jedna próbka
z Estonii. • •
• •
• • R1a1a7b L260 - gałąź P - polska /
zachodniosłowiańska |
Drzewo
|
Według niektórych przypuszczeń powstanie mutacji M420, a zatem i haplogrupy R1a, dokonało się gdzieś na stepach
europejskiej Rosji lub Ukrainy, na północ od Morza Kaspijskiego lub
Czarnego, jako w refugium podczas ostatniego zlodowacenia. Tego refugium nauka
jednak nie potwierdziła. Bardziej zasadne będzie wskazywać to
miejsce w okolicach gór Ałtaju, gdzie wśród niektórych plemion
(dziś językowo ałtajskich, tureckich i rosyjskich)
występuje ojcowska haplogrupa R1-M173 do 20%, czyli w stosunkowo najwyższym
zagęszczeniu.
Inna istotna sprawa - to powstanie późniejszej mutacji M17, czyli wielkiej haplogrupy R1a1a, łączącej wielkie populacje Europy, Azji
Środkowej i Indii, czyli w skrócie - haplogrupy eurazyjskiej.
Według niedawnej pracy
zespołu hinduskich badaczy (ostatnio S.Sharma i zesp., styczeń 2009)
ojcowska dla R1a1a haplogrupa R1a1-SRY1532.2 miałaby się wyłonić w
północnych Indiach na terenie Kaszmiru; albo w środkowych Indiach (w
plemieniu Saharia) około 18.400 lat temu (licząc 25 lat na
pokolenie). Tam wśród plemion Kashmiri Pandits zidentyfikowano 9 osób (4%
spośród badanych) z tą haplogrupą, a w plemieniu Madhya Pradesh
Saharia 13 osób (23% badanych). Według tych badań, na terenie
Kaszmiru lub Indii miałaby powstać także następna mutacja, M17, dająca początek haplogrupie R1a1a. Dała ona zarazem początek - według tych
autorów - hinduskim Indoeuropejczykom, głównej populacji
Indii. Obecnie do indoeuropejskiej populacji
R1a1a w Indiach należy do 16% hinduskiego społeczeństwa. I co
najciekawsze, ta głównie populacji utworzyła
najwyższą kastę hinduską, kapłańskich braminów
albo przynajmniej nią liczebnie zawładnęła .
Przeciętnie blisko połowa, a w głównej ich grupie - nawet 72%
kapłanów-braminów wyróżnia się hg R1a1, podkreślają badacze
hinduscy. Daty jakoby sugerują, że z hinduskiej populacji
R1a1a miałaby wywodzić się późniejsze populacje
Słowian i innych pokrewnych ludów. Autorzy zamieszczają daty tej
haplogrupy na Bliskim Wschodzie i w Europie - 11.200 lat temu, a w Azji Centralnej
- ok. 8600 lat temu.
(Inne badania wskazują
także na Iran jako miejsce pochodzenia hg R1a1a (praca M. Requeiro i
zesp. 2006; badania niedokładne!). Tam haplogrupę R1a1 (SRY1532.2)
zidentyfikowano ogólnie w 3% badanych, a w północnym Iranie - nawet 15%).
Jednak
A. KLOSOW, częściowo wspierany badaniami innych genetyków, wskazuje
"kolebkę" R1a1a ogólnie w południowej Syberii, w
pobliżu Chin. Oto
fakty:
|
Alan Bittles et al. 2007 R1a1a
wywodzi się z południowej Syberii; mutacja M17 powstała tam
20.000 lat temu bowiem na taki czas
wskazują jej haplotypy STR, które Klosov w swojej pracy
opublikuje. |
Ostatnią
wielką populacją, wydzieloną z eurazyjskiej haplogrupy R1a1a,
są Ariowie, którzy dali początek językowo indoeuropejskiej
części społeczności Iranu i Indii (w Indiach 16%, czyli
około 100 milionów). Jednak znacznej części tej populacji Klosow
przypisuje bezpośrednie pochodzenie z południowej Syberii lub Chin.
Mianowicie, prof. A. Klosow,
przeprowadzając rewizję dotychczasowego datowania haplogrup
europejskich i azjatyckich, w tym także R1a1a, zauważył, że
dokonane przez historyków, językoznawców i archeologów datowanie inwazji półwyspu
Indyjskiego przez Ariów z północy i zaistnienie tam indoeuropejskiego
języka dopiero na połowę drugiego tysiąclecia p.n.e. jest
zgodne tylko z poprawnym datowaniem i genealogią hinduskiego haplotypu
R1a1a, wyraźnie identycznego z haplotypem wschodniosłowiańskim,
lecz nieco młodszego.
Bowiem spośród
Prasłowian wschodnich, których wspólny przodek wywędrował 2750
lat p.n.e na ukraińsko-rosyjskie stepy, część populacji,
którą datuje sie na 2300 lat p.n.e, migrowała dalej na wschód, na
teren dzisiejszego północnego Kazachstanu i południowego Uralu,
tworząc tam rozległą kulturę andronowską (z centrum
nad Jeziorem Aralskim). Niewielka grupa z tej populacji zbudowała sobie
około 1800 r. p.n.e m.in. miasto/zamek o nazwie Arkaim na południowym
krańcu Uralu. Został on jednak porzucony po około 200 latach.
Bowiem po jakiejś katastrofie, którą można kojarzyć z
wybuchem (około
1628 lat p.n.e)
wulkanu Santorini (zwanym też Thera) na Morzu Egejskim i który
pochłonął cywilizacje minojską, a na szerokiej przestrzeni
północnej półkuli na długi czas zakłócił pogodę
(znaczne ciemności i obniżenie temperatury) ludność
Arkaim z R1a1a porzuciła gród i udała sie na południe,
zaludniając Kirgizję i Tadżykistan (tam dziś wśród
ludności - do kilkadziesiąt procent genetycznych ex-Słowian
R1a1a!) oraz północne Indie, a chyba osobno - północny Iran (badania
irańskie są dotąd niewystarczające).
Co do
hinduskiej populacji R1a1a trzeba wiedzieć i brać pod uwagę,
że na podstawie tekstów hinduskiej świętej księgi Rygwedy i
innych badań, zwłaszcza archeologii i językoznawstwa, światowa nauka
dotąd uważa indoeuropejski lud Ariów, który dał początek
kastom braminów hinduskich i hinduskim Indoeuropejczykom, raczej za
najeźdźców z północy w połowie II tysiąclecia p.n.e.,
co potwierdza genealogia genetyczna. Dlatego Genetyka na portalu Eupedia
podkreśla: "Kapłańska kasta braminów składa
się niemal wyłącznie z haplogrup R1a1, R2 i J2a (chociaż
R1a1a stanowi dwie trzecie z linii), przybyłych w ramach indoaryjskiej
migracji z zewnątrz w epoce brązu 3500 lat temu". Genealogia
Y-DNA w badaniach A.K. zdaje się potwierdzać pochodzenie
językowo indoeuropejskich Ariów od Słowian wschodnich, o czym
niżej, w rozdziale o ekspansji Słowian.
[Inne
mutacje w haplogrupie R1a1a-M17, oprócz R-M458, zostały rozpoznane
poza tradycyjnymi ziemiami słowiańskimi, zwłaszcza w Azji;
stąd tam kolejne haplogrupy: R1a1a1-M56, R1a1a2-M15, R1a1a3-M64.2,
R1a1a4-P98, R1a1a5-PK.5, R1a1a6-M434, a ostatnio - podgrupa polskiej:
R1a1a7a. Te wszystkie są nieliczne, co najwyżej kilkuprocentowe
lub tylko śladowe i bardzo młode. Spodziewane jest wykrycie nowej
mutacji wśród obecnych R1a1a w Skandynawii razem z Kirgizją oraz
kilku mutacji w R1a1a w Indiach].
Kolejne
badania A. KLOSOWA
21.11.2009 r. Journal of Gnetic Genealog 5/2 2009.
opublikował właśnie ważną dwuczęściową
pracę A. Klyosova na temat rozpoznania interesujących nas
haplotypów w haplogrupie R1a1a, datowania pierwszych wspólnych przodków i migracji. Odnotowuje też i krótko
omawia rezultaty badań Underhilla et al. 2009.
DNA Genealogy, Mutation Rates..., cz. I
DNA Genealogy, Mutation Rates..., cz. II
W części 1. autor omawia swoje sposoby
rozróżniania haplotypów i trzy metody obliczania czasu poszczególnych
haplotypów od wspólnego przodka (TMRCA);.stosowane razem, sprawdzają
się wzajemnie i dają znaczną dokładność i
pewność.
W
części 2.
znajdują się praktyczne zastosowania tych metod do obliczania czasu
głównych populacji haplogrupy R1a1a (M17) (bez szczegółowego
zajmowania się dopiero co wykrytą podgrupą R1a1a7) oraz innymi
wybranymi haplogrupami i populacjami
|
Oto
haplogrupa R1a1a - wg. Anatole Klyosov 2009 AZJA |
Inne wybrane haplogrupy: 16.000 lat - Haplogrupa R1b (M343) - geneza tej
haplogrupy |
Badania innych autorów
Archeolodzy zaś,
np. David Anthony (zob. Dyskusja n.12
i mapy), jako pośrednią
praojczyznę Słowian przed ich europejską migracją
wskazują teren starej kultury Boh-Dniestr, oraz horyzont Jamna
(kurhanowy), a z którymi trzeba jakoś połączyć także
kulturę ceramiki sznurowej (Corded Ware). To wszystko na wielkim terenie
od Karpat Wschodnich po Ural.
Najnowsze badania, ogłoszone przez Underhilla (4.11.2009; zob. niżej,
rozdz.6) jednak nie bardzo wskazują na ten wschodni teren, lecz na ziemie
polskie. Potwierdza go Klosow i Rożanskij (7.12.2009; zob. niżej,
rozdz.6)
Zadziwiająca jest jednolitość, liczebność, a zarazem
rozległość na mapie Eurazji ludności z haplogrupą R1a1a-M17 i jej podgrupą R1a1a7-M458: od
Uralu na wschodzie, po linię Łaby i wschodnich Alp na zachodzie; od
Bałtyku, a nawet wybrzeży Norwegii i zachodniej Szkocji na
północy, po Indie, Macedonię i Morze Czarne na południu!
Największą częstotliwość tej haplogrupy w Europie, do
ponad 60% ludności, stwierdza się w pasie między Bałtykiem
a Morzem Czarnym, obejmującym całą Polskę i Ukrainę.
|
|
Haplogrupa R1a1a
(M17) Widoczne zagęszczenia w rejonie Polski, Ukrainy i Rosji,
północnego Pakistanu i Indii oraz Azji Środkowej (Kirgizja). |
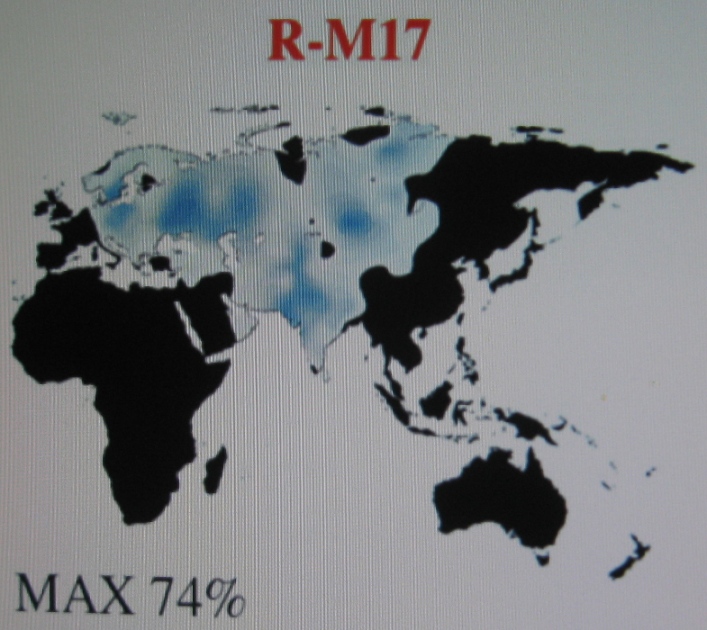
|
|
Ludność Europy w starożytności na początku nowej ery według portalu Eupedia.tree2. Zob. rozmieszczenie R1a na wschodzie, R1b na zachodzie, I2ana południu i wschodzie, I1, R1a, R1b na północy nordyckiej,N na północy rosyjsko-fińskiej i uralskiej, E w rejonie Grecji. |
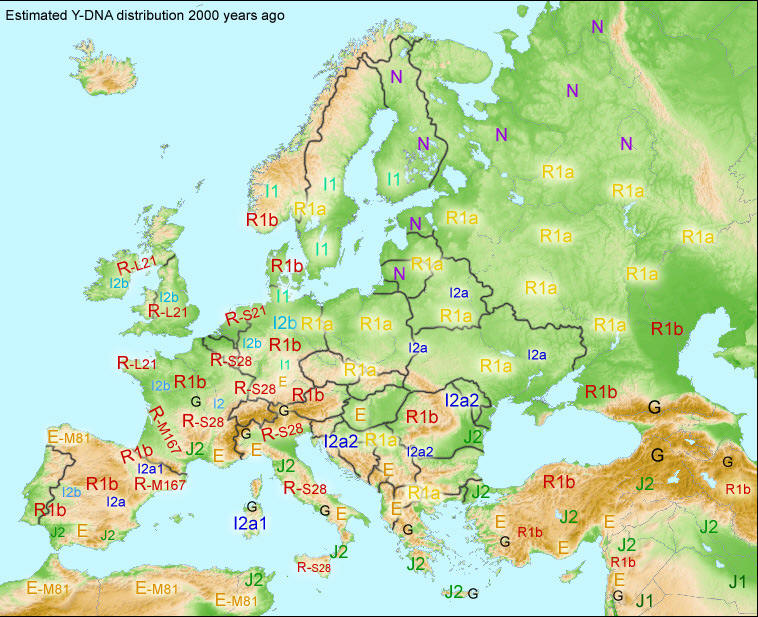
Haplogrupą
R1a1a wyróżniają się, jak widzimy, szeroko rozprzestrzenione na
kontynencie europejskim ludy słowiańskie i bałtyjskie oraz te,
które się dowodnie z ich wspólnoty wydzieliły (łącznie z
Kirgizami, Tadżykami i indoeuropejskimi Ariami/Hindusami, o czym
niżej). Mając na uwadze to, że spośród kulturowych
wyróżników etnosu język jest najbardziej trwałym dobrem, a
według niezmiennej zasady, zakłócanej tylko nadzwyczajnymi
zdarzeniami,
język
otrzymuje się w rodzinie od przodków i praprzodków jako podstawowe dobro
tradycji rodowej i plemiennej,
więc początek słowiańskiego języka wolno i trzeba
łączyć z początkiem nie tylko polskiego oddziału
R-M458, ale i euroazjatyckiego R-M17, a nadto jego substratu trzeba
dopatrywać się w ojcowskiej haplogrupie R1a1 (gdzieś w Azji
Środkowej), a nawet jeszcze wcześniej - w haplogrupie R1
(tamże?, Azja Zachodnia?), w której języki satemowe hg R1a
łączyły się, jako w jednym źródle, z kentumowymi
językami hg R1b i wszystkich jej podgrup (zob. niżej).
Rozwój europejskiej POPULACJI R1a1a i R1a1a7. SŁOWIANIE
Populacja R-M17, obecna na Bałkanach od dziesiątego tysiąclecia
przed Chr. (ale nie jej późniejsza podgrupa R-M458) na pewno
uczestniczyły jakoś w procesach rewolucji neolitycznej od
około 5500 lat przed Chrystusem: w powstaniu i upowszechnianiu się
rolnictwa, którego skutkiem był szybki rozwój demograficzny i migracje
ludności. Wynikiem tych migracji był udział R1a1a w
powstaniu rozmaitych kultur archeologicznych nad Dunajem, np. ceramiki wstęgowej,
oraz na północ i wschód od Karpat, np. kręgu ceramiki sznurowej z
kulturami grobów jednostkowych i amfor kulistych. Nad Dniestrem Bohem i
Dnieprem dochodzi do symbiozy z ostatnia fazą kultury Cucuteni-Trypolie u
wschodnich Karpat, od rzek Prut i Seret do Dniepru. Następuje teraz
rozwój kultury Prasłowian dalej w kierunku wschodnim, ku Wołdze i
Uralowi, powstanie około 4000 lat przed Chr. kurhanowego kręgu
kulturowego Jamna. Na stepach pontyjsko-kaspijskich słowiańskim
kulturom trzeba przypisać udomowienie konia, stosowanie powozu i upowszechnianie
hodowli zwierząt domowych (zob. Renfrew, Archeologia i język, PWN
2001, s. 249)
Te
kultury były wynikiem pierwszej ekspansji R1a1a ku Środkowej i
Wschodniej Europie; z tego terenu dokonała się około 3000 lat
przed Chr. także dalsza migracja w różne rejony Europy,
zwłaszcza na zachód i wschód. Czas i kierunek wędrówek jest dziś
rozpoznawany przez warianty haplotypów w STR w haplogrupie R1a1a, zakodowane w
Y-DNA dzisiejszych mieszkańców Europy lub w znalezionych kopalnych
szczątkach praprzodków, choć rzadko (bowiem niektóre nowsze plemiona
Prasłowian paliły swoich zmarłych; zresztą nie tylko
Prasłowianie).
Genealogia Y-DNA (A.Klosow) ustaliła, że przodek dzisiejszych
wschodnich Słowian z hg R1a1a (M17) żył na ziemiach Ukrainy i Rosji
od około 2750 lat
przed Chr,
(Uwaga. Nie
znaleziono dotąd w rejonie Ukrainy i Rosji oczekiwanych przez naukowców
genetycznych śladów starszej ludności R1a1a, zwłaszcza takich,
których by można datować na czasy ukraińskiego refugium podczas LGM,
którego domyślał się już O.Semino i zesp. 2000, tak jak je
znaleziono na Bałkanach. Ukraina nie jest więc pierwotną
kolebką wszystkich Słowian. Zwłaszcza nie wolno ich genezy
datować tam na czasy niedługo przed V wiekiem po Chr., jak to
głoszą niektórzy archeolodzy polscy, tzw. skrajni allochtoniści
("dnieprowcy") np. K. Godłowski, a za nimi także niektórzy
genetycy (zob. Dyskusja n.5), chyba wprowadzeni w
błąd publikacjami znanego niemieckiego archeologa i rasisty Gustawa
Kossiny (+1931). Około VI-VII w. po Chr., a więc już w czasach
historycznych, ówcześni Słowianie z Ukrainy swoją
reemigracją tylko może nieco zasilili osadnictwo południowych
Słowian na Bałkanach, przerzedzone klęskami w wojnach z
cesarstwem wschodnim oraz na terenie Polski i krajów sąsiednich).
Dziś na Ukrainie, Białorusi i w Rosji żyje około 50% ludności
naszych haplogrup (w niektórych starych miastach do 70%). Genealogia Y-DNA
prowadzona przez A. Klosowa zidentyfikowała nawet starożytnych
założycieli dziewięciu plemion ruskich z czasu około 2000
lat p.n.e i późniejszego, nazwanych imiennie dopiero we
wczesnośredniowiecznych zabytkach pisanych, m.in. bliscy nam
Wołynianie, Bużanie i Biali Chorwaci (kilka kopalnych szczątków
ludzi owego osadnictwa hg R1a1 zidentyfikowano nawet w Krasnojarsku w
południowej Syberii).
Równie stare jak na Ukrainie i Rosji, bo z około 2700 lat p.n.e. korzenie
plemion R1a1a (M17) znajduje sie na terenie Niemiec, a więc jest tam daleko
wcześniejsze niż geneza Germanów, zwłaszcza tam, gdzie dziś
nadal żyje skondensowane osadnictwo także słowiańskiej hg
R1a1a7 Serbołużyczan (choć zwykło się je
datować dopiero na wiek VII po Chr.). Potwierdza to np. niedawne, dokonane
w 2005 r., odkrycie cmentarzyska kilkunastu zmarłych, a wśród
nich trzech męskich osób z hg R1a1a w Eulau, 40 km za Lipskiem nad
rzeką Soławą (niem. Saale) w dorzeczy Łaby w Saksonii. Owe
groby ofiar jakiejś starożytnej przemocy pochodzą z około 2600
lat p.n.e i
wyróżniają się świadectwami bogatej kultury życia
rodzinnego i małżeństwa egzogamicznego. Plemię to
prezentowało kulturowe środowisko tzw. ceramiki
sznurowej, kojarzonej powszechnie z Indo-Europejczykami. Wśród
ludności niemieckiej do dziś około 10% wykazuje
haplogrupę R1a1, których wspólny przodek żył tam już
około 2700 lat p.n.e.
Czyżby owi nadłabscy Serbowie, a zwłaszcza ich bałkańscy
protoplaści, zanotowani przez tzw. geografa bawarskiego jako Zeriuani
(Serbianie?), byli tymi, "z których wszystkie plemiona
słowiańskie powstały i ród swój - jak zapewniają -
wywodzą"?
Można ich
śmiało identyfikować z tzw. kulturą grobów jednostkowych,
która pojawia się około 2700 przed Chr. w ramach paneuropejskiej
kultury ceramiki sznurowej. Jej południowy region rozciągał
się od Małopolski przez Śląsk i Czechy po Solawę
(dopływ Łaby), z Serbołużyckim Eulau. Północny
region tej kultury przylegał do Bałtyku i rozciągał
się od Kujaw i dolnej Wisły, przez Wielkopolskę, rejon dolnej
Odry i Meklemburgię po Dolną Łabę (z tym horyzontem
można kojarzyć późniejsze R1a1a w jaskini Lichtenesteinhohle z
ok. 1000 przed Chr.), zob. Dyskusja, n.16.
Także słynna kultura łużycka może być bez
zastrzeżeń przypisana głównie (choć nie tylko)
słowiańskiej hg R1a1a. Populacje haplogrupy R1a1a i jej podgrupy
żyją dziś w Polsce w największym w całej Europie
jednorodnym zagęszczeniu, bo blisko
60% ludności
Polski.
|
Na tablicy rozmieszczenie
haplogrup w dzisiejszych krajach europejskich; zob. R1a1 w czwartej
kolumnie. Liczby oznaczają procenty owej haplogrupy w poszczególnych
krajach. R1b - to zachodnioeuropejska. |
Nieco później niż w Polsce R1a1a (bez uwzględnienia
podziału na R1a1a i R1a1a7!) pojawia się w Czechach(dziś 30%), Słowacji (40%), Węgier (32%) i Anglii (5% mieszkańców), Irlandii (dziś m.in. klany Donaldów i Douglasów),
w Szkocji; w Norwegii (do 30% mieszkańców części
zachodniej). Stąd osadnictwo R1a1 emigrowało
doIslandii (dziś
tam 23% ludności). Nadto do Austrii - dziś ok. 25%. Także w Szwecji i Norwegii tj. w nadmorskich regionach
zachodnich, zwłaszcza wśród przybyłych z Rusi dawnych wikingów;
nie dziwi więc udział wikingów w tworzeniu słowiańskiego
państwa ruskiego w IX-X w.), Na Litwie, Łotwie i Estonii dziś odpowiednio 38, 40 i 32%.
W sumie, na mapie geno-geografii Europy największe zagęszczenie
słowiańskiej hg R1a1a ciągnie się szerokim pasem od
Bałtyku po Morze Czarne (co jest wodą na młyn tym, którym marzy
się Polska "od morza do morza"!).
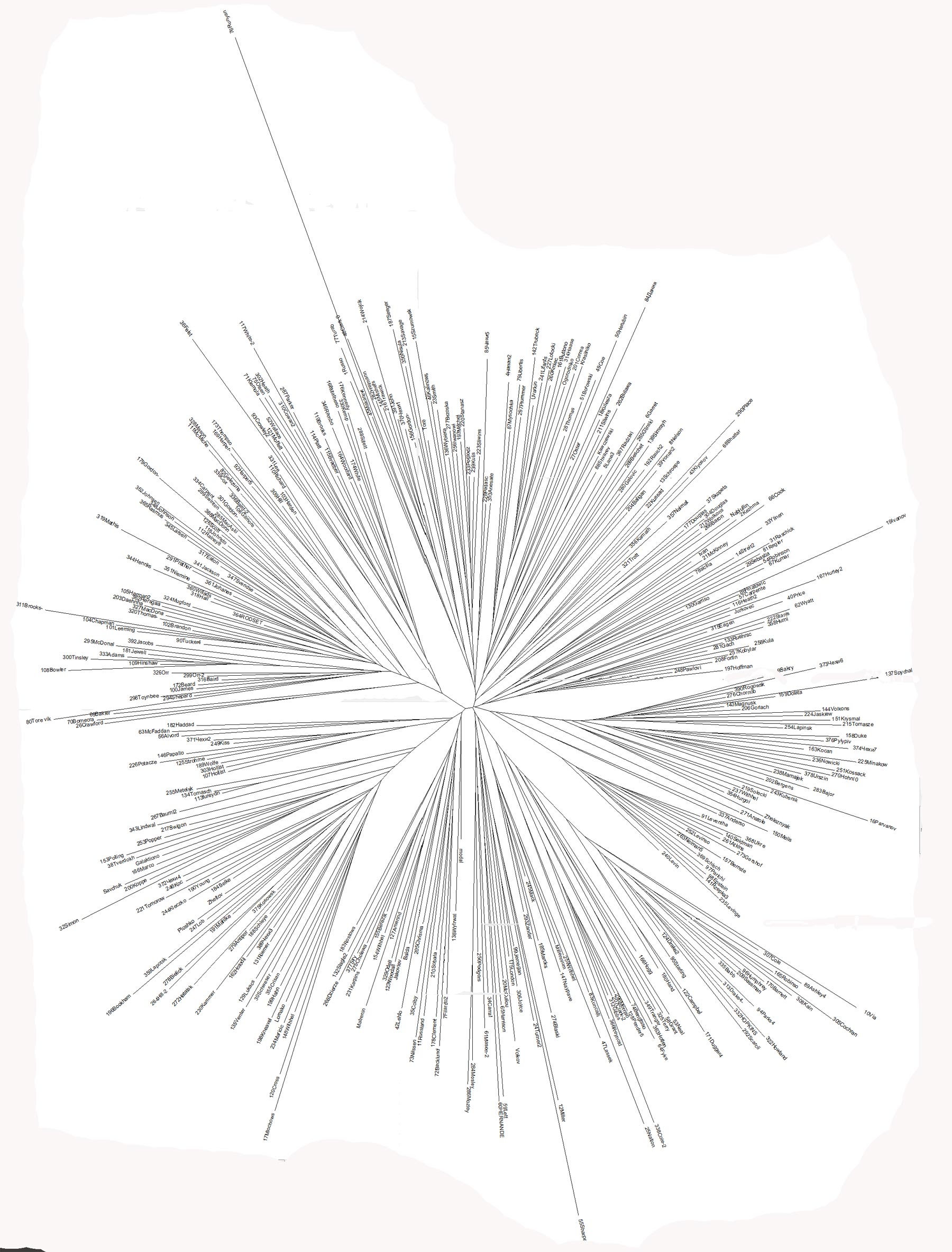
Drzewo
genealogiczne europejskiego oddziału R1a1a.
Gałęzie haplotypów, datowanie i ich rozmieszczenie na mapie
Europy
ŻRÓDŁA
1. I. Rożanskij i A. Klosow, listopad 12009
- kompendium aktualnej wiedzy o europejskiej części populacji
R1a,
(gałęzie przed rozpoznaniem mutacji M458 i haplogrupy R1a1a7)
"Haplogrupa R1a: haplotypy, linie genealogiczne, historia, geografia"
И. Рожанский и А. Клёсов, Гаплогруппа R1a: гаплотипы, генеалогические линии, история, география,
w: Вестник Российской Академии ДНК-генеалогии t.2, nr.6 (listopad), 2009 r., s. 974-1099.
2. A. Klosow i I. Rożanskij,
grudzień 2009
o haplogrupie R1a1a7, czyli gałęzi środkowoeuropejskiej i
zachodniosłowiańskiej
"Podgrupa R1a1a7 - M458 - populacje, geografia, historia"
w: Вестник Российской Академии ДНК-генеалогии t.2, nr 7 (grudzień), 2009 r.,
s.1200-1216.
w języku rosyjskim, gruntowny komentarz do pracy Underhilla et al. 2009
(zob. niżej) w temacie haplogrupy R1a1a7-M458,
uzupełnienie pracy Rożanskiego-Klosowa
"Haplogrupa R1a: haplotypy, linie genealogiczne, historia, geografia" Вестник t.2, nr 6, 2009 r. (zob.
wyżej).
3. I. Rożanskij i A. Klosow,
styczeń 2010
o bliższym genealogicznym związku gałęzi
skandynawskich z kirgiską.
"Migracje z południowej Syberii i Azji Środkowej z punktu
widzenia genealogii Y-DNA"
Вестник Российской Академии ДНК-генеалогии, tom
3, nr 1, 2010 r.
4. I. Rożanskij - 2009-2010
kilkanaście map rozmieszczenia gałęzi haplotypów R1a1a1, R1a1a2,
R1a1a3 i R1a1a7
Google-mapy;
5. P. Szwarew, Forum Rodstvo.ru,
omówienie map i drzew genealogicznych
Гаплогруппа R1a: гаплотипы, генеалогические линии, история, география, И. Рожанский и А. Клёсов
6. I Rożanskij, A. Klosow,
styczeń 2010
"Migracje z południowej Syberii i Azji Środkowej do
północnej Europy z punktu widzenia genealogii Y-DNA"
Миграции из южной Сибири и Средней Азии в северную Европу с точки зрения ДНК-генеалогии
w: Вестник Российской Академии ДНК-генеалогии Том 3,
№ 1 2010 январь
http://www.lulu.com/items/volume_67/8049000/8049755/2/print/8049755.pdf
Igor Rożanskij, Schematyczne
drzewo genealogiczne europejskiej R1a1a i podgrup
(zaktualizowana: 30.03.2010)
|
|
Drzewo genealogiczne Nazwy
gałęzi kolejno (od lewej) 2. Środkowoeuropejska R1a1a7 3. Zachodniosłowiańska R1a1a7 i 7b 5. Centralna eurazyjska-3 - niebieska
8. Мłodsza skandynawska
9. Centralna eurazyjska-1 -
żółta 10. Północnokarpacka 12. Centralna eurazyjska-2 -
czerwona 13. Kirgiska i młodsza 16. Północnozachodnia 17. Stare europejskie: podziałka skali czasu 1000 lat Źródło obrazu: Rodstvo.ru |
|
Objaśnienia 12000 lat temu - oddzielona
haplogrupa R1a1a2 z mutacją M157 i haplogrupa R1a1a3 z mutacjami
M64.2/87/204, zwane Stare europejskie. 4800 lat temu (2800 przed Chr.)
- oddzielone osiem populacji, choć także w warunkach "szyjki
butelki", w tym datowanej na ten czas populacji R1a1a* na tzw. Ruskiej
Równinie, zapewne jednej (nr 1) z trzech gałęzi Centralnej
eurazyjskiej. Haplotypy tych trzech gałęzi żyją na
szerokim terenie od Irlandii przez Polskę po Półwysep Indyjski.
Rozwój więc tych gałęzi trzeba datować na czas daleko
przed migracją indoeuropejskich Ariów spod Uralu do Indii i Iranu. Dzisiaj żyjące 8 najstarszych populacji powstały w
latach 1500-2000 przed Chr. Ich przodkowie
podlegali więc długo sytuacji "szyjki butelki". Uwaga.
Gałąź młodsza skandynawska ze względu na pochodzenie
jest na mapie łączona z kirgiską.
Źródło obrazu: http://s56.radikal.ru/i151/1004/e7/b8c82e15bbcc.jpg |
|
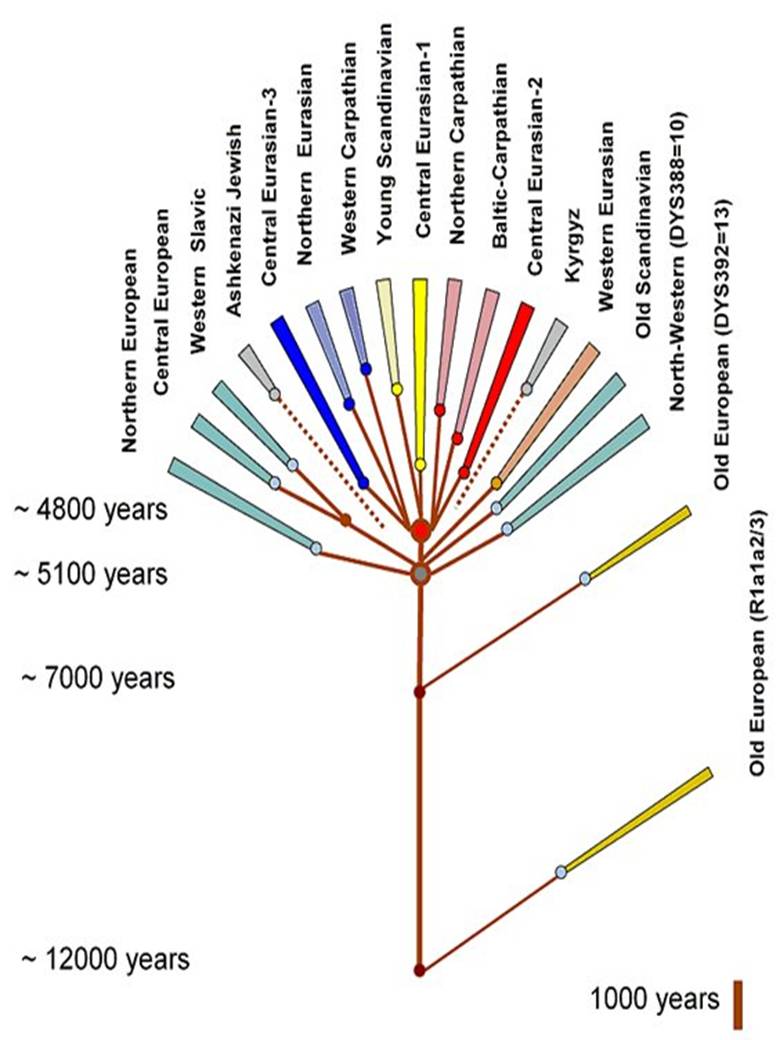
Podział, datowanie i mapy gałęzi
europejskich populacji haplogrup R1a1a i podgrup
wg Igora Rożanskiego 2009 (wybór)
Wszystkie mapy zob.: Google-mapy; zob. też forum Rodstwo.ru
|
|
R1a1a-M17/198 |
|
|
|
|
|
|
|
To hg. R1a1a7; ich wspólny przodek: 1900 p.n.e. |
||||
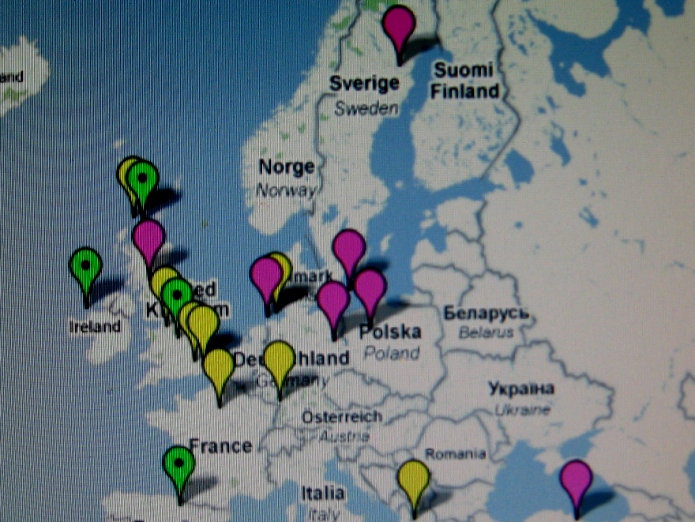
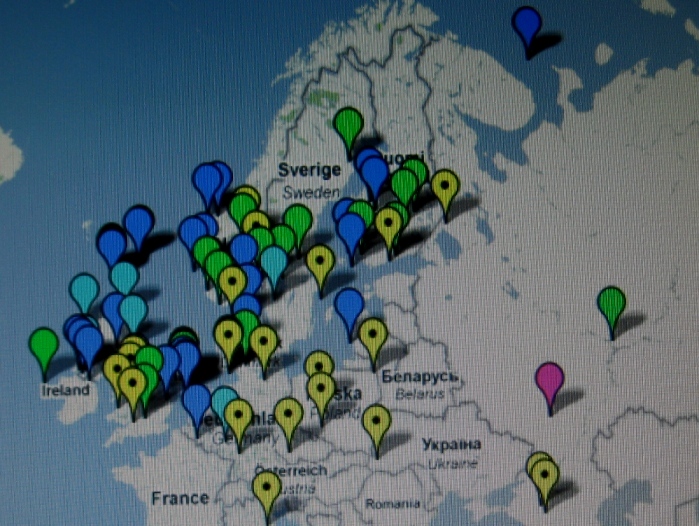
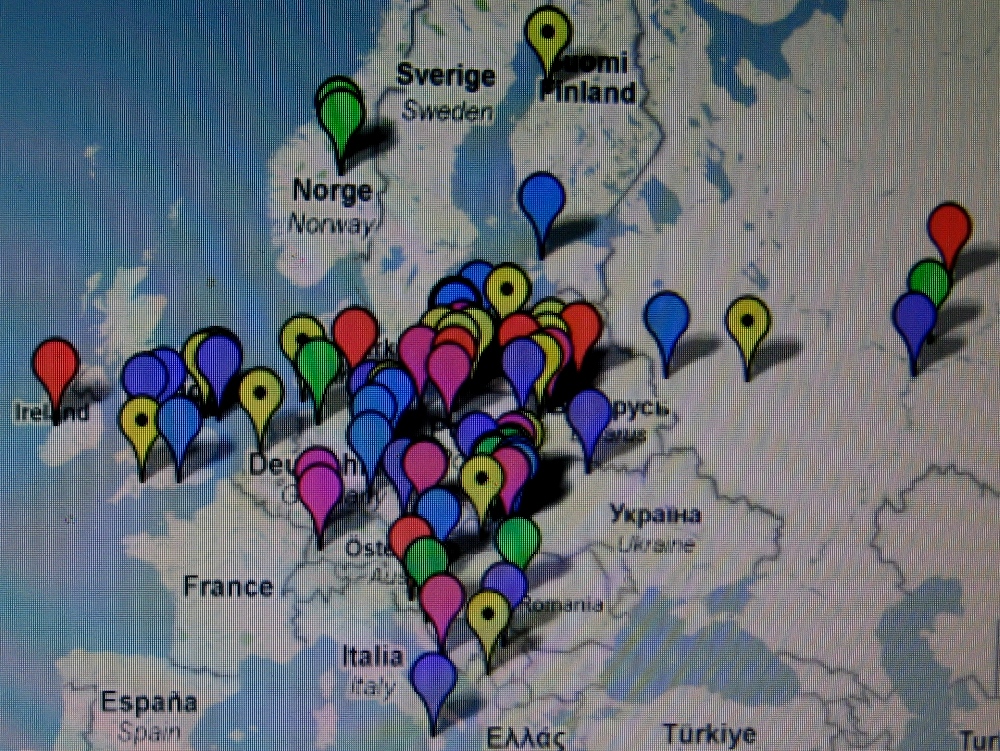
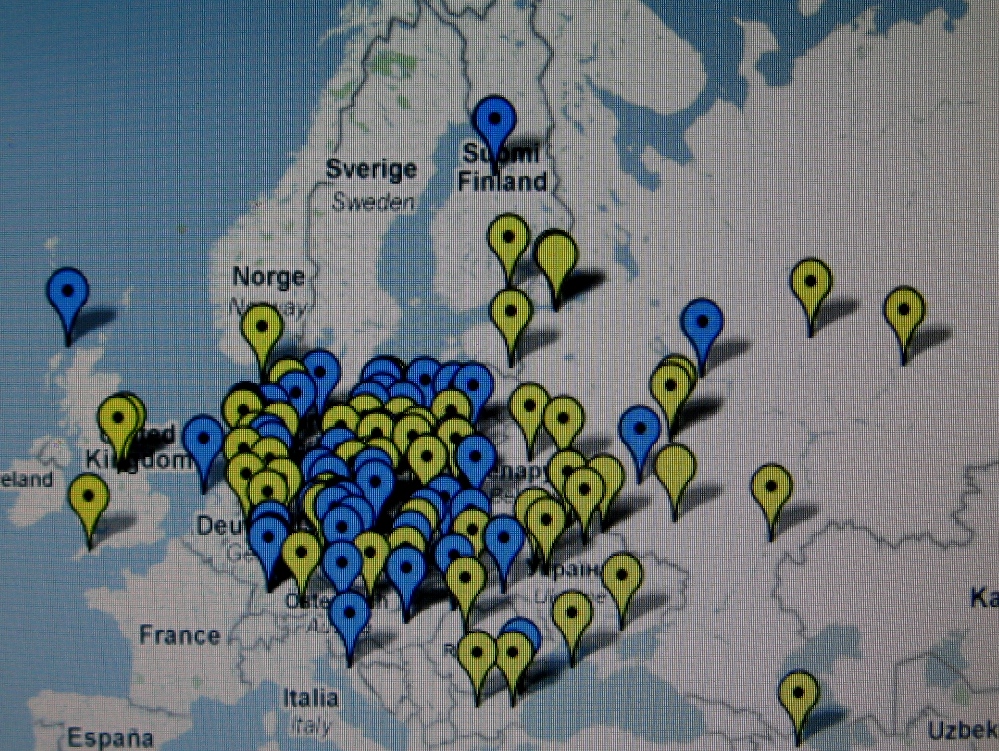
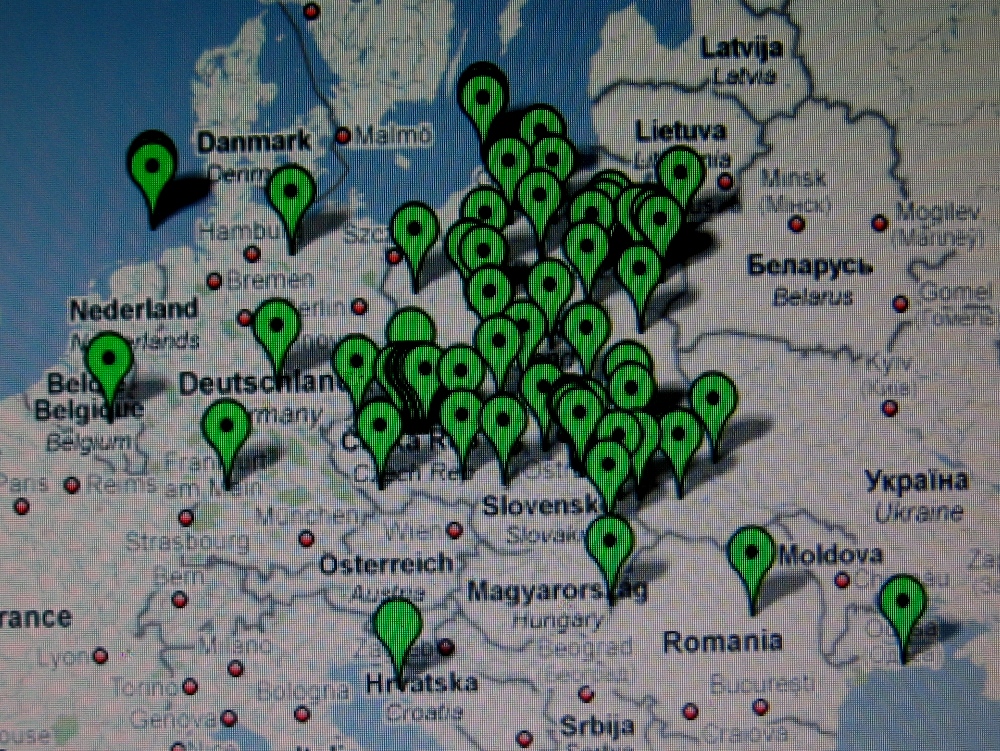
Te i pozostałe mapy I. Rożanskiego z Ukrainy zdają
się nie
potwierdzać tezy
niektórych archeologów o naddnieprzańskim
pochodzeniu Polaków i ich haplogrupy R1a, i to dopiero w V wieku po Chr. Co więcej,
wyraźnie zauważa się, że Polska leży w centrum
starożytnego osadnictwa słowiańskiego haplogrupy R1a.
W Rosji natomiast o
rezultatach pobierania próbek DNA wprost pisze się:
Газета.ru: "Поскреби русского - найдешь поляка"
'Poskrob Ruskiego - znajdziesz Polaka' (14.01.2008).
|
|
Drzewo
genealogiczne http://www.rodstvo.ru/forum/index.php?showtopic=37&view=findpost&p=12572 Sporządził:
Павел Шварёв (Rodstvo.ru) |
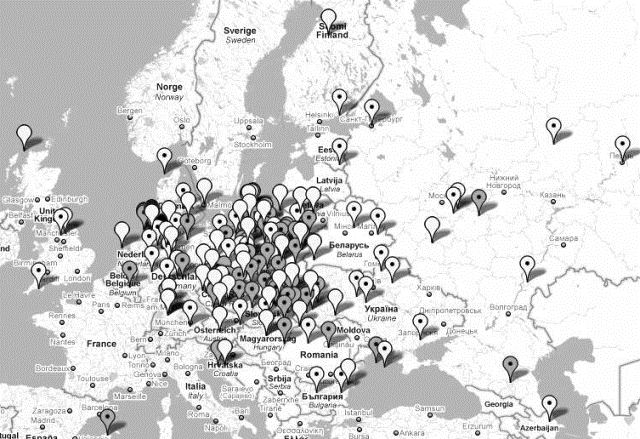
Najstarsze haplotypy R1a1a-M17 w Europie
zostały w 2008 r. zidentyfikowane przez Klosowa wśród ludności
niektórych krajów bałkańskich: Serbia, Bośnia, Kosowo i
Macedonia, w materiale, który zebrali i opublikowali M. Perićić i
L.Barać. A. Klosow datuje je na około dziesiąte tysiąclecie
przed Chr. (około 9600 r.).
Zespół genetyków P. Underhill et al.
(listopad 2009, zob. niżej) wykrył zaś ważną
europejską mutację R-M458, tworzącą nową
haplogrupę R1a1a7, datując ją, z grubą przesadą, na
dziewiąte tysiąclecie przed Chr. i (dość trafnie)
wskazując na rejon Polski jakby na teren jej powstania i jakby na
praojczyznę znacznej populacji Słowian środkowoeuropejskich,
zwłaszcza polskich. Underhill zidentyfikował też
dwie inne haplogrupy, R1a1a6 i R1a1a7a. ale
niewielkiego znaczenia.
ROŻANSKIJ i KLOSOW w swoich obszernych publikacjach
dokonują uporządkowania i genealogicznego opracowania obecnej wiedzy o
populacjach haplogrupy R1a1a i podgrup R1a1a2, R1a1a3 i R1a1a7 oraz ich
historii w Europie i Eurazji. Autorzy mają aktualnie do dyspozycji
około 2000 haplotypów z bazy YSearch i geograficznych projektów FTDNA oraz
z laboratoriów rosyjskich i prywatnych, najwięcej 25-markerowych.
Prowadzą w tej chwili jedyne takie prace w świecie.
A. Najdawniejsze gałęzie populacji R1a1a w Europie (od
około 10000 lat przed Chr.)
(po modernizacji drzewa i datowania 30.03.2010 przez Igora
Rożańskiego)
- trzy gałęzie w postaci trzech starych haplogrup: R1a1a (zob.
wyżej, drzewo: gałąź środkowa, dolna
część), R1a1a2 wraz z R1a1a3 (gałąź dolna po
lewej) oraz wydzielona seria z DYS392=13 lub14 (gałąź dolna po
prawej). Wspólny przodek tych trzech gałęzi żył
około 8-10 tysięcy lat przed Chr.
1. Haplogrupa R1a1a1-M17/198. Ta
haplogrupa jest jakby na osi ewolucji całej haplogrupy R1a1a i jej
podgrup. Datowana jest w Europie na około 12000 lat temu ((10000 lat przed
Chr.) Jej dzisiejszym etapem rozwoju są trzy gałęzie centralnoeurazyjskie.
2. Stara europejska I (Haplogrupy
R1a1a2 i R1a1a3). Powstanie populacji haplogrup R1a1a2 i R1a1a3 datowane
jest na lata około 2400 przed Chr., zapewne na nowym miejscu, po migracji
z pierwotnej ojczyzny w południowowschodniej Europie. Są nieliczne i
praktycznie nierozdzielne. Wspólny przodek ich i Centralnej eurazyjskiej
gałęzi żył około 10000 lat przed Chr. Jest to zapewne
zarazem czas powstania ich mutacji
(Przy okazji informacja: Kolejne
haplogrupy R1a1a4 i R1a1a5 są puste - nie znaleziono osób, a odkrytą
przez Underhilla haplogrupę R1a1a6 reprezentuje tylko kilkanaście
osób w Pakistanie i Omanie).
3. Stara Europejska II (Oddział z DYS392=13); datować
go trzeba na 125 i 175 lat przed Chr., w zależności od haplotypów
25 czy 37 markerowych; jego wspólny z gałęzią
środkowoeurazyjską przodek żył około 5100 lat przed
Chr. Jest charakterystyczne, że grupa DYS392=13 z oddziałów
R1a1a2 i -a3 ma też trzy haplotypy spośród trzynastu w starej
bałkańskiej gałęzi z 9 tysiąclecia przed Chr.
Zob.
wyżej mapa 1, Stara europejska gałąź R1a1a.
Niesprzyjające warunki bytowe (tzw. szyjka butelki) lub dryf populacji
zdziesiątkowały ich liczebność i nie pozwoliły im na
pełny rozwój demograficzny. Zanim jednak do tego kryzysu doszło,
prawdopodobnie około polowy trzeciego tysiąclecia przed Chr., mieli
zapewne wydatny wpływ na rozwój archeologicznych kultur naddunajskich,
środkowoeuropejskich i dalszych. Archeogenetyka zapewne będzie
miała szansę wypowiedzieć się na ten temat, jak to się
stało już odnośnie szkieletów z Eulau, Lichtenstein-Hohle,
Krasnojarsk itd.
B. Młodsze gałęzie R1a1a (od
około 3100 lat przed Chr.)
Nazwy
gałęzi są tylko orientacyjne, ustalone arbitralnie.
Gałęzie, które otrzymały orientacyjną nazwę
"europejskie" (sześć), wydzielają się
wyraziście, a "eurazyjskie" (drugie sześć) -
wydzielające się mniej wyraziście. Pięć z nich, a
właściwie sześć (dzieląc R1a1a7 na dwie
gałęzie) oddzieliło sie od osi R1a1a około 3100 lat przed
Chr.
4. Północno-zachodnia, tzw.
dziesiętniki (z
DYS388=10) - głównie na Wyspach Brytyjskich i w Norwegii. Wspólny przodek
ich linii starszej żył około 925 lat przed Chr., a linii
młodszej - 575 po Chr. Wspólny przodek ich obydwu linii żył
około 1700 lat przed Chr., natomiast wydzielenie się mutacji
DYS388=10 z populacji R1a1a nastąpiło 3100 lat przed Chr.
5. Starsza skandynawska,
której MRCA datowany jest na około 2100 przed Chr.. Do niej dopisano, jako
ojcowską, populację z Eulau nad Solawą w Niemczech (zob. I.Rożanskiego mapa
gałęzi starszej skandynawskiej), datowaną
izotopowo na rok około 2600 lat przed Chr., nosicieli przypisywanej
Prasłowianom kultury ceramiki sznurowej. Wyróżnia ją szczególnie
mutacja markera YCAII=19,21, podczas gdy w Europie wynosi ona przeważnie
19-23. Oprócz tego marker DYS565=13.
Skandynawska gałąz
R1a1a zapewne wniosły elementy języka prasłowiańskiego i
indoeuropejskiego (odmiany satemowej) do tworzącego się etnosu
germańskiego (zob. niżej, w rozdz. 9).
6. Zachodnioeurazyjska, rozproszona niemal po całej
Europie. Jej MRCA można datować na 1400 lat przed Chr.
7. Północnoeuropejska, skoncentrowana jest wyraźnie nad
Dolną Wisłą. Jej dwie podgałęzie datowane są na
lata 1700 i 250 przed Chr., a ich wspólny przodek żył około 2000
lat przed Chr.
8.
Środkowoeuropejska R1a1a7, wyróżnia
się własną mutacją -M458.
Obecna jest w krajach Europy Środkowej, głównie w trójkącie od
Czech do ukraińskiego Zakarpacia i Basenu Dunaju (także
Małopolski i Śląska, a nawet Wielkopolski, kraju Polan); jest
liczna - ponad 20 milionów mężczyzn w Europie. Wyłoniła
się prawdopodobnie na południe od Karpat, zapewne w Basenie
Dunaju. Wyróżniają ją rzadkie markery DYS464e-f-g, a
zwłaszcza mutacja M-458, wspólna z gałęzią
zachodniosłowiańską. Jej przodek żył około roku 500 przed Chr. ,
a wspólny przodek z Zachodniosłowiańską - 1900 lat przed
Chr.
W dunajsko-karpackiej
ojczyźnie tej gałęzi mógł się zrodzić jakiś
"kult" cesarza Karola niemieckiego i stąd upowszechnić
się ogólnosłowiańskie słowo "król". W
dunajsko-karpackiej ojczyźnie mogła być też praojczyzna
Polan polskich i naddnieprzańskich (zob. u Nestora). Interesujące
jest i to, że ekspansja części tej populacji (zwł. z
DYS464e-f-g), zapewne w połowie I tysiąclecia n.e., szła - jak
wskazuje drzewo genealogiczne - w kierunku z
zachodu na wschód, na Ruską Równinę i ku Morzu
Białemu, więc w przeciwnym kierunku niż inne, znane w
tradycyjnej literaturze naukowej, migracje w czasie wędrówek ludów - ze
wschodu na zachód (s.1026).
(Tak
więc ten jak i inne fakty nie potwierdzają tezy skrajnych allochtonistów
polskich o nieobecności Słowian na naszych ziemiach przed
połową pierwszego tysiąclecia naszej ery i o jedynym tylko
kierunku ich ekspansji: znad Dniepru ku Wiśle i Odrze!).
9. Zachodniosłowiańska
R1a1a7b. Wyróżnia
się mutacją L260,
podporządkowaną mutacji M-458 gałęzi
środkowoeuropejskiej R1a1a7, wydzielona z niej około 1900 lat przed Chr.
Rozmieszczona jest głównie w Polsce, Czechach, Słowacji i na Rusi
Zakarpackiej. Powstała zapewne nieco na północ od Karpat.
Stanowią ją populacje głównie w dorzeczach Wisły, Odry i
Łaby (Polska, Czechy i Słowacja).
Ich wspólny przodek żył w latach około 575 przed Chr.
Obydwie gałęzie dość długo żyły w pewnej od
siebie separacji, skoro Środkowoeuropejska gałąź
migrowała na wschód i północ raczej sama.
UWAGA 1. Gałęzie
Zachodniosłowiańska i Środkowoeuropejska, mające wspólna
mutację M458 z około 3100 lat przed Chr., oddzieliły się
wtedy od gałęzi Centralnej eurazyjskiej (CEA); znalazły
się jednak w sytuacji długo trwającej demograficznej "szyjki
butelki" i rozwinęły się dopiero w połowie
pierwszego tysiąclecia przed Chr.
UWAGA
2. Na ojczystym terenie gałęzi Środkowoeuropejskiej i
Zachodniosłowiańskiej, a właściwie zapewne już
wśród ich dalekich przodków R1a1a na Bałkanach, dokonała sie
satemizacja języka dotąd kentumowego oraz formowanie języka
praindosłowiańskiego, a potem oddzielonego praslowiańskiego.
Ostatecznie ludności
R1a1a, R1a1a7 i R1a1a7b, żyjącej na na niewielkim terenie
Europy Południowo-Wschodniej czy Środkowo-Wschodniej należy
przypisać dokonanie podstawowych prasłowiańskich innowacji
językowych, których zaistnienie po migracji części tej populacji
na wschód i rozproszeniu ludności byłoby już niemożliwe.
C. Haplogrupa R1a1a od około 2800 lat przed Chr.
10.11.12.
Trzy gałęzie Centralne eurazyjskie.
To
trzy duże populacje, powiązane wspólnym przodkiem z około 2700-2800 lat przed Chr.,
upowszechnione są od Wysp Brytyjskich, poprzez Europę
kontynentalną i rejon tzw. Ruskiej Równiny, do Indii (zob. mapa Rożanskiego,gałęzie
rozróżnione kolorami). Wspólny przodek gałęzi żółtej
datowany jest na około 1875, niebieskiej - 1700 i czerwonej - 1575
lat przed Chr.
Po około 2800 r. przed Chr. dokonało się przybycie pierwszych
niewielkich populacji CEA z naddunajskiej Europy na Ruską Równinę (i
dalej, na teren całej "postsowieckiej" Rosji). Haplotypy modalne
trzech gałęzi Centralnej eurazyjskiej, jak i ich MRCA, są prawie
identyczne. Wschodnim ich populacjom R1a1a można przypisać m.in.
wielkie wschodnioeuropejskie kultury archeologiczne (horyzont) Jamna, Poltavka,
grobów zrębowych i wielką kulturę Andronowo ze słynnym
"grodem" Arkaim (1800 lat przed Chr). Przedłużeniem ich
ekspansji była migracja Ariów ku Azji Centralnej, Iranowi i Indiom.
Ekspansja tych populacji prawdopodobnie zasilała także w pierwszym
tysiącleciu p.n.e. inne, zwłaszcza środkowoeuropejskie
populacje, które wcześniej przeżywały demograficzną
"szyjkę butelki". Zapewne z rejonu nadwołżańskiej
i nadkaspijskiej Rosji dokonała się ekspansja R1a1a ku Kirgizji i
Skandynawii (młodsza gałąź z około połowy
pierwszego tysiąclecia naszej ery).
Gałęzi Centralnej
eurazyjskiej (CEA) na pewno przysługuje nazwa "Satemowcy",
czyli lud języka indoeuropejskiego odmiany satem (od satem 'sto' w językach
słowiańskich i indoirańskich; w odróżnieniu od centum'sto' w
językach zachodnioeuropejskich).
Zob. osobny
opis tej gałęzi
Centralnej eurazyjskiej na
stronie genealogii rodzinnej.
13. Północnoeurazyjska,
młoda, rozpowszechniona zwłaszcza na północny-wschód od Polski:
Białoruś, Ukraina i Rosja. Jej MRCA datowany jest na około
przełomu ery starej i nowej.
14. Zachodniokarpacka,
niewielka i bardzo rozproszona. Jej MRCA żył około 1675 lat
temu, a wspólny jej przodek z gałęzią środkowoeuropejską
żył około 2700 lat przed Chr.
15.
Młodsza skandynawska -
obecna także na Wyspach Brytyjskich i Islandii. Składa się z
trzech podgałęzi, różniących sie od starej skandynawskiej w
YCAII=19,21 i DYS565=12. Datowane są na lata 200 przed Chr. do 700 po Chr. Z nimi wiąże się też
wikingów skandynawskich, a także słynny klan MacDonalda z około
1325 roku. Zarówno podobieństwo haplotypu, jak i dane archeologiczne i
tradycje kulturalne (sagi skandynawskie i kult Odyna) wskazują na
pokrewieństwo tej populacji skandynawskiej ze środkowoazjatyckimi,
kirgiskimi. Czas ekspansji kojarzy się z okresem wędrówek ludów
około V-VI wieku (brała w niej udział zapewne także
populacja haplogrupy Q z południowej Syberii, obecna do dziś w
Skandynawii). Gówna ojczyzną ekspansji mógł być teren na
północ od Morza Kaspijskiego.
16. Północnokarpacka,
rozpoznana głównie na terenie Polski. Jej MRCA datowany jest na 550 lat
przed Chr.
17. Bałto-karpacka,
mało wyraziście oddzielona i skoncentrowana. Jej trzy podgrupy
datowane są na czas od około 750 lat przed Chr. do
300 po Chr. Zaś ich wspólny przodek żył 1000 lat przed Chr.
18.
Kirgiska, obecnie złączona na mapie z
gałęzią młodszą skandynawską. Wspólny
przodek gałęzi kirgiskiej z młodymi skandynawskimi żył
około 800-1000 lat przed Chr. Migracja ku Kirgizji mogła dokonać
się w czasach tzw. wędrówek ludów, czyli około połowy I
tysiąclecia n.e.
19.
Gałąź żydów aszkenazyjskich, rozpowszechniona
zwłaszcza w pasie między wschodnim Bałtykiem a
północno-zachodnim brzegiem Morza Czarnego. Ta populacja, mająca
swojego przodka na 900 rok n.e., zapewnie w Niemczech, miała z
całą haplogrupą R1a1a wspólnego przodka (ale wówczas jeszcze
nieżydowskiego) około 4100 lat przed Chr.
UWAGA.
W rejonie Europy
Środkowej nastąpiło spotkanie ludności hg. R1a1a z
ludnością kultury pucharów lejkowatych i amfor kulistych (zapewne hg.
I1 i I2), oraz zainicjowanie i/lub upowszechnianie kilku innych kultur,
zwłaszcza ceramiki sznurowej (Eulau), toporów gładzonych i grobów
jednostkowych. Regres demograficzny, trwający do początku pierwszego
tysiąclecia przed Chr. w rejonie Europy Środkowej nie był
całkowity i nie oznaczał katastrofy kulturowej, zwłaszcza
że na sąsiednich ziemiach dzisiejszej Ukrainy i Rosji Europejskiej
tego regresu nie było, było więc jakieś okresowe wsparcie i
demograficzne i kulturowe dla ziem rejonu Środkowej Europy
|
|
Oto drzewo haplotypów hg R1a1a7*(
Klosow) z danych Underhilla. |
|
|
Mapa zagęszczenia populacji haplogrupy R1a1a7 w
Europie Środkowej. |
Dla porównania -
datowanie siostrzanej haplogrupy R1b w Europie, nowa publikacja Klosowa, Gaplogruppa
R1,
s. 249-299, zwł. tablica na s. 292-298 (uważać na rachubę
czasu u Klosowa: YBP, czyli "od obecnego czasu").
Вестник
Российской
Академии
ДНК-генеалогии,
Wiestnik t.3, nr 2, luty 2010.
* * * * *
Inne badania europejskiej i polskiej części haplogrupy R1a1a i R1a1a7
1) Gwozdz i Mayka.
Z wynikami pracy A. Klosowa w dużym stopniu harmonizują badania
amerykańskich genealogów polskiego pochodzenia, Piotra Gwoździa
(Peter Gwozdz) i Wawrzyńca Majki (Lawrence Mayka).
L. Mayka prowadzi program FTDNA Polish
Project .
Rejestruje tam się wyniki analiz przeprowadzonych w pracowniach
FamilyTreeDNA dotyczących materiałów z DNA, dostarczonych przez
Polaków ; internetowa strona przedstawia rezultaty badań i identyfikacje
haplogrup Polaków (aktualnie, w lutym 2010 - 1202 osób), a w haplogrupie R1a1a
- podział na klastry, w zależności od haplotypów STR.
Dokładnym rozpracowaniem polskich haplotypów z haplogrup R1a1a i R1a1a7
zajmuje się P. Gwozdz.
Na
swojej internetowej stronie Polish clades prowadzi analizę haplotypów,
grupuje, oblicza czas populacji i ich migracji. W opublikowanej w JOGG Journal of
Gnetic Genealog 5/2 2009 jego
pracy i pojawia się więcej informacji na temat metody
pracy i jej rezultatów (część
II). Czas wspólnego przodka haplotypu P (u
Klosowa: gałąź zachodniosłowiańska) z haplogrupy
R1a1a7 Gwozdz datuje ogólnie na pierwsze tysiąclecie przed Chr. Ekspansje
tej populacji z ziem polskich datuje na lata 400-500 po Chr. W innym miejscu
pierwszego przodka tego haplotypu datuje na 400 lat po Chr.
Słowiański haplotyp K, odpowiadający haplogrupie R1a1a,
łączy nieśmiało z "kulturą" kurhanów (4000
lat przed Chr.), ale dopuszcza i wcześniejsze tysiąclecia, jak to
zakładał Underhill.
Dwie populacje z haplogrupą
R1a1a*
Dwie populacje z haplogrupą R1a1a7.
|
Haplogr. R1a1a, typ
K |
Haplogr. R1a1a, typ
KB |
Haplogrupa
R1a1a7 Wspólny przodek (por. datowanie
przez Klosowa: 1900 lat, |
Haplogr. R1a1a7, typ N |
Haplogr. R1a1a7, typ P |
|
1) Typ K i typ K-Borderline to dwa klastry starszej
haplogrupy R1a1a w Polsce i krajach słowiańskich, z których
wywodzą się typy N i P. Obok typu K i KB wraz z ich podtypami,
aktualnie wymienia się także klaster E i "mój" klaster F. 2) Typ N w pracy Gwozdzia odpowiada gałęzi
środkowoeuropejskiej u Klosowa; a typ P - gałęzi
zachodniosłowiańskiej. Autorzy przypuszczają, że dalsze
badania ujawnią mutacje w SNP, co pozwoli powyższe
typy/gałęzie nazwać osobnymi haplogrupami. 3) Wewnątrz zbioru R, czyli
"pozostałych" (około 20% całej polskiej R1a) oraz
zbioru U, czyli jeszcze nieoznaczonych (też znaczny procent),
niektóre haplotypy wydają się starsze od typu K, mogą
więc należeć do najwcześniejszych w Europie. 4) Odmiennym niż polski i kontynentalny jest typ
L. Jest to typ R1a1a skandynawski, północnoeuropejski
(wikingowie?): np. Norwegia, Wyspy Brytyjskie, Polska (mały procent) i Kirgizja.
Razem to 23% całej R1a w bazie Ysearch. 5) Powyższe mapy
uwzględniają tylko tych, którzy są zarejestrowani w
PolishProject FT-DNA, admin. L.Mayka, a statystyka z PolishProject oraz bazy R1a w
Ysearch. Dane te będą stale
ulegać zmianie i rozrastać się. |
||||
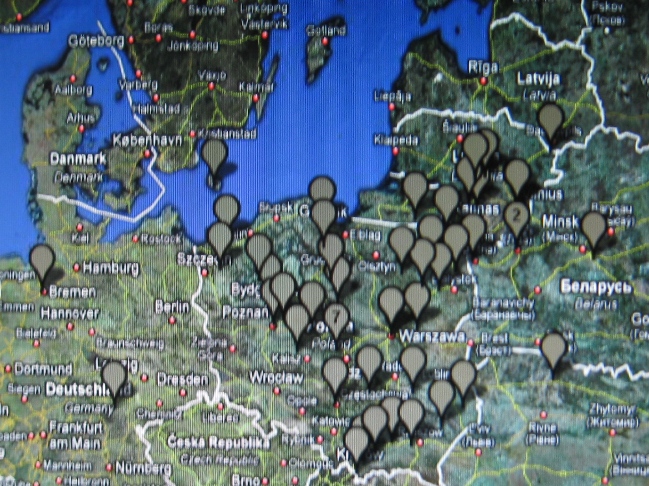
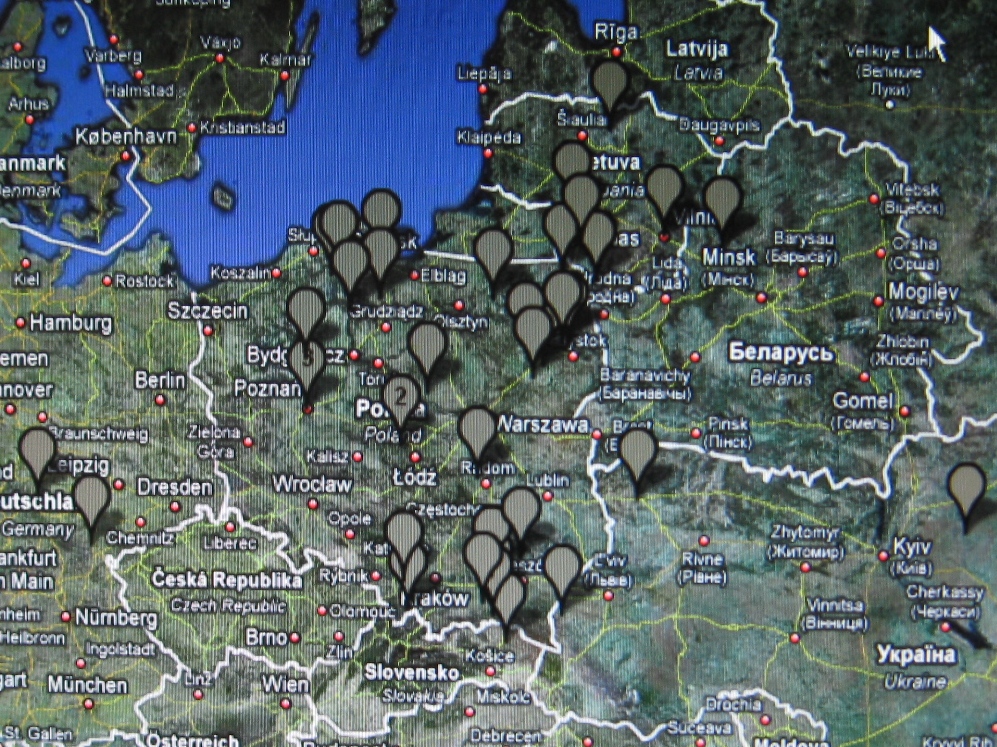
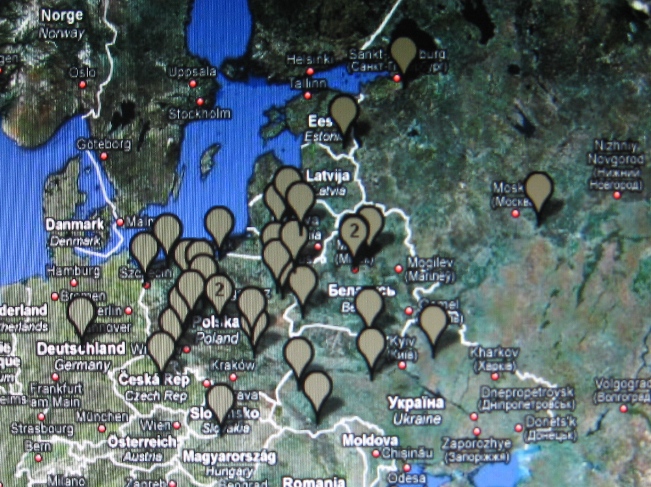
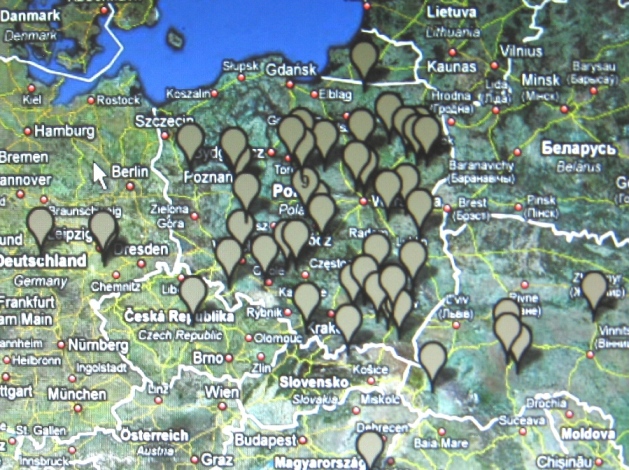
W marcu 2010 r. Mayka i Gwozdz wydzielili w haplogrupie R1a1a kilka klastrtów,
w tym F i H, które odpowiadają gałęziom centralnoeurazyjskim w
pracach rosyjskich genealogów.
2) Badania Petera Underhilla
Publikacja Underhill et al.
(publikacja 4.11.2009), znacznie zmieniła dane wcześniejszych
autorów, wskazały na Polskę jako na praojczyznę
Słowian i miejsce powstanie polskiej mutacji R-M458 dla
słowiańskiej populacji Europy. Oto niektóre dane polskie i wybranych krajów.
Czas wspólnego przodka podano w tysiącach lat temu, obliczony jest
według stawek Zhivotovsky'ego, a więc chyba niewłaściwie
obliczonych i dwu-trzykrotnie zawyżonych.
Oto kilka danych Underhilla odnośnie (nieprawidłowego!)
datowania populacji R1a1a-M17 i R1a1a7 -M458
w kilku krajach w tys. lat:
Polska 11300 i 10700; Słowacja 11200 i 8300; Czechy - 5700; Niemcy 9900 i
7500
Rosja europejska 8700 i 8000;
Ukraina 7400 i 4700; Indie 14000; Pakistan 15000
|
Na lewo: mapa
zagęszczenia, czasu haplogrupy R1a1a (M17) i drzewo genealogiczne
grupy i podgrup. Na prawo: mapa
zagęszczenia, datowania i związku z kulturami archeologicznymi populacji
R1a1a7 (M458) - według: Underhill et al. 2009. |

Zob. praktyczne zastosowanie
genealogii Y-DNA haplogrupy R1a1a na rodzinnej stronie Pietrzakowie.
UDZIAŁ SŁOWIAN w TWORZENIU STAROŻYTNYCH
KULTUR
Populacje haplogrupy R1a z jej podgrupami są wraz z populacjami haplogrupy
R1b głównymi twórcami nie tylko języków z rodziny indoeuropejskiej,
ale także wielkiego dziedzictwa kultur duchowych i materialnych
świata starożytnego i nowożytnego, zwłaszcza Europy.
Ubogi i prymitywny materiał ceramiczny tzw. kultury praskiej, korczakowskiej
i podobnych, przypisywany przez archeologów wzorem G. Kossiny Słowianom VI
w. n.e. i następnych, już teraz, w świetle wiedzy o haplogrupach
R1a1a i R1a1a7, już nie niepokoi. Ta kulturowa prostota mogła
być wytworem tylko niektórych, do V w. zapewne jakoś odseparowanych,
niewielkich populacji słowiańskich, poruszonych ówczesnymi migracjami
i zmianami demograficznymi; nie może ona więc świadczyć o
prymitywizmie całej olbrzymiej populacji słowiańskiej
wcześniej i w połowie pierwszego tysiąclecia po Chrystusie,
obecnej przecież w mniejszym lub większościowym procencie na
terytorium od Pirenejów, Islandii, wybrzeży Norwegii (Wikingowie) i
Bałkanów - po Ural i daleko dalej na wschód, twórców wielu wielkich,
znanych i wyrafinowanych kultur.
Ci właśnie,
żyjący na terenach Europy Środkowej i Południowo-Wschodniej
Prasłowianie R1a1a i R1a1a7 (oraz populacje z nich
wydzielone) byli na tych ziemiach twórcami lub współtwórcami wielu kultur
archeologicznych, jak to ukazują mapy na portalu
Eupedii. Zob. Dyskusje
18
|
Główne archeologiczne kultury,
Tu trzeba z niewielkimi zmianami powtórzyć to, co wyżej powiedziano
o populacji R1b, że zdefiniowaną genetycznie populację
R1a trzeba łączyć z satemowym wariantem języka
indoeuropejskiego, pewnym zubożeniem form ceramiki na rzecz ceramiki
sznurowej, początkiem epoki brązu w Europie, obniżeniem
kultury rolnej na rzecz hodowli zwierząt i pasterstwa, przejściem
od matriarchatu do patriarchatu, wprowadzeniem grobów jednostkowych i
kurhanów nagrobnych, udomowieniem i wykorzystaniem konia oraz rozwojem cech
wojennych. |
Uwzględniwszy
dotychczasowe dane antropologii, archeologii i językoznawstwa
historycznego, wielu może odczuwać ogromne zaskoczenie dostarczonymi
przez genealogię genetyczną, zwłaszcza geno-geografię
danymi i powyższymi wnioskami. Ale trzeba też zachować
ostrożność, przynajmniej do czasu przebadania ludzkiego
materiału archeologicznego oraz udoskonalenia metod datowania, które
nadal doprowadzają do wielu rozbieżności pozostawiają wiele
niepewności chronologicznych.
W każdym razie teraz od wielu archeologów, zwłaszcza skrajnych
słowiańskich allochtonistów, powyższe fakty będą
wymagały wiele przysłowiowej wielkoduszności i przyznania
się do pomyłek, zrozumiałych przecież z powodu dotychczasowej
niewystarczalności narzędzi do ustalania etnicznego i czasowego
znakowania kultur.
* * * * *
W nowym i korzystnym dla nas świetle staje także wyjątkowa zwartość i
liczebność etniczna oraz tożsamość
słowiańska Polaków, obecnych od wielu tysięcy lat na tej ziemi (Underhill
ustalił nawet blisko 12 tysięcy lat), choć może z okresami
znacznego regresu demograficznego, tzw. pustki osadniczej. Widać to
szczególnie na tle sąsiednich krajów, zwłaszcza Niemiec (gdzie
Germanów z normańskiej haplogruopy I-1 - nieco ponad 40%, ze zgermanizowanej
części celtyckiej hg R1b 40%, a ze słowiańskiej R1a1a
i R1a1a7 - 20%) oraz mieszkańców Czech i Słowacji (gdzie oprócz
Słowian żyje dość liczna grupa celtycko-germańska z
R1b odpowiednio Czechy 28% i Słowacja 17% oraz hg
południowoeuropejska I-2, po 18%). I zadziwia nadto, zwłaszcza
wobec genetycznej różnorodności w haplogrupie
zachodnioeuropejskiej, genetyczna jednolitość całej
haplogrupy słowiańskiej R1a1a, w której dotąd nie doszło do
żadnej dalszej mutacji i wydzielenia się osobnej podgrupy genowej; na
kontynencie europejskim (Polacy, Słowacy, Macedończycy,
Ukraińcy, Rosjanie...) różnimy się tylko osobniczymi allelami w
STR.
Szkoda tylko, że ci Słowianie, choć tak liczni, nie utworzyli
wcześniej, już w starożytności, jednej zwartej grupy
politycznej i własnego wielkiego i trwałego państwa. Teraz
trzeba ich identyfikować nie tylko z późniejszymiSklawinami,
Antami i Wenedami (odVI w. po Chr.), ale i pod wieloma
innymi, wcześniejszymi nazwami ludów starożytnej Europy,
może łącznie z niektórymi "Sarmatami" (od III w.
przed Chr. po późne średniowiecze), niektórymi "Scytami"
(znanymi co najmniej od VII w. przed Chr.), Aorsami, Neurami, Budynami, Melanchlajnami, a
zwłaszcza Ariami (ok. 1500 lat przed p.n.e.). Trzeba ich
więc widzieć, i to już w starożytności, na
Bałkanach i nad Dunajem, na rozległym terenie od Bałtyku po
Morze Czarne, od ziem Rusi po Morze Aralskie, a nawet od Uralu po ziemie
północnych Indii, Pakistanu i Iranu - wszędzie tworzących
niekiedy prymitywne, ale niekiedy i bardzo rozwinięte i wyrafinowane
kultury archeologiczne (zob. wyżej i na mapach Eupedii:
http://www.eupedia.com/europe/neolithic_europe_map.shtml) oraz
systemy protopaństwowe.
Dzisiejsza przestrzenność i niemal jednolitość języków
słowiańskich, nakładająca się na
przestrzenność haplogrup R1a, każe nam zakładać
podobną przestrzenność wcześniejszego,
niezróżnicowanego jeszcze języka prasłowiańskiego.
Biorąc jednak pod uwagę fakt, że język jest głównym
czynnikiem różnicującym ludzi i wytwarzającym
świadomość odrębności etnicznej, ta wielka jednolitość
i przestrzenność prajęzyka była zarazem hamulcem procesów
politycznych. To tłumaczy trudności w identyfikacji i niewielką
obecność Słowian w tekstach starożytnych historyków i
geografów. Brak wyraźnego czynnika zagrożeń i między
etnicznej rywalizacji nie wpływał na przyspieszenie integracji
społecznej i mobilizacji gospodarczej. Dopiero w bliższym
zetknięciu się z południowo i zachodnioeuropejskimi,
zorganizowanymi społecznościami i państwami Słowianie
uświadamiają sobie swoją odrębność, a nawet
nadają sobie nazwę etniczną: Słowianie, jako ci
mówiący zrozumiałymi słowami, w przeciwieństwie do
"niemówiących" (psł. "niem") lub
mówiących niezrozumiałą mową, czyli Niemcami. Nazwą
tą określano początkowo zapewne prawie wszystkich
Nie-Słowian.
8.
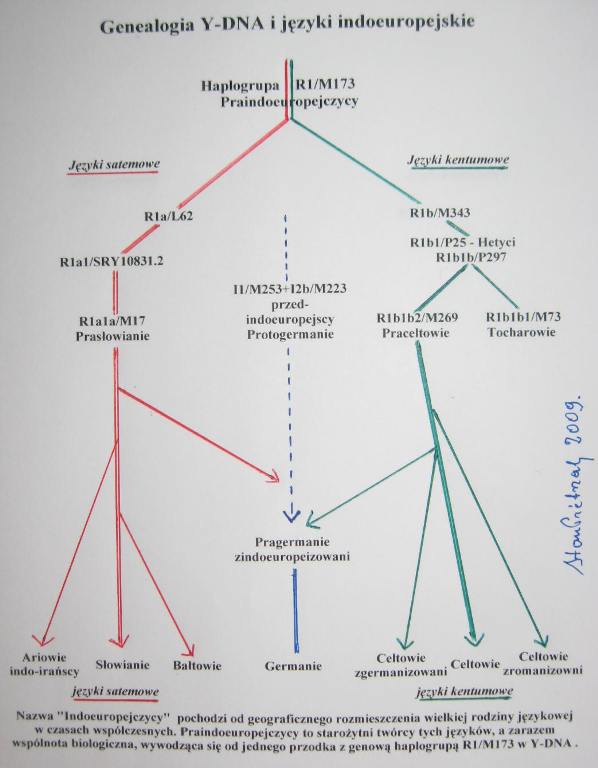
GENEZA JĘZYKÓW INDO-EUROPEJSKICH.
Satem i kentum w świetle genealogii Y-DNA
Wyraźne podobieństwo języków prawie w całej Europie i
głównego języka na Półwyspie Indyjskim skłoniło
językoznawców do uznania ich pokrewieństwa i nadania im nazwy
"języki indoeuropejskie". Te związki językowe są
świadectwem istnienia u podłoża jednego języka,
wytworzonego i używanego przez jedną wspólnotę ludzką,
żyjącą w zwartej grupie na stosunkowo niewielkim obszarze.
Językowi temu nadano nazwę "praindoeuropejski", a jego
twórców i użytkowników nazwano "Praindoeuropejczykami".
Według dość
powszechnie przyjętego w językoznawstwie poglądu prawidła
ewolucji języków (glottochronologia) miałyby wskazywać na
trzecie tysiąclecie przed Chr. jako na czas istnienia jeszcze
jednego i niepodzielonego języka.
Gdyby chodziło o
ojczyznę języka i wspólnoty prandoeuropejskiej to analiza
najstarszego słownictwa, wspólnego dla wszystkich języków
indoeuropejskich, miałaby wskazywać na stepy południowej Ukrainy
lub Rosji (choć nie brak uczonych wskazujących na Azję Środkową,
Anatolię w Azji Mniejszej lub na Bałkany).
Podział języka
praindoeuropejskiego na znane nam dziś języki od krańców Europy
po Półwysep Indyjski nastąpił na skutek rozprzestrzeniania
się i migracji ludności oraz dokonywanych innowacji
językowych w nowym małych grupach, od siebie oddalonych i
narażonych na kontakty z zupełnie obcymi językami.
Główny jednak
podział na grupę satemową i kentumową miałby być,
według wielu językoznawców, rezultatem innowacji w tych
językach, które pozostawały w centrum obszaru języków indoeuropejskich
(j. satemowe, zwł. słowiańskie i indoirańskie) oraz braku
innowacji w tych językach, które z powodu migracji znalazły się
na peryferiach tego obszaru (j. kentumowe, zwł. celtyckie, romańskie
i germańskie).
1. Cechą pierwotnego języka praindoeuropejskiego była
palatalność spółgłosek tylnojęzykowych, czyli
zmiękczonych k', g', g'h.
2. Po podzieleniu i
rozejściu się wspólnoty w jednej jej części
doszło do spirantyzacji owych spółgłosek tylnojęzykowych
palatalnych, czyli przeszły one w spółgłoski szczelinowe typu ś, sz, s. Na skutek tego np. słowok'entum przybrało brzmienie satem (j. awestyjski) lub sto (j. słowiańskie) i stąd
nazwa tej grupy języków: satemowa.
3. W drugiej zaś
części pierwotnej wspólnoty językowej doszło tylko do
odmiękczenia (dyspatatalizacji) spółgłosek pierwotnie
palatalnych. Na skutek tego np. słowo k'entum 'sto' przyjęło brzmienie kentum 'sto' i stąd nazwa tej grupy
języków: kentumowa.
Do niedawna uważano, że do językowej innowacji w postaci
satemizacji doszło w centrum pierwotnego terytorium językowego; a ta
innowacja nie dotarła do nadal kentumowej peryferii. Jednak teraz
językoznawstwo ma już doskonalsze narzędzie,
umożliwiające dokładniejsze ustalenie warunków i
okoliczności powstania wspólnoty języków indoeuropejskich oraz
fundamentu podziału. Tym narzędziem jest genealogia Y-DNA. Ona bowiem
wskazuje na istnienie konkretnej, biologicznej i pierwotnej wspólnoty
praindoeuropejskiej w postaci populacji haplogrupy R1. Obecnie łatwo dostrzec,
że językami kentumowymi wyróżniają się przede
wszystkim populacje haplogrupy R1b (celtyckie, romańskie i
germańskie). Natomiast językami satemowymi wyróżniają
się populacje haplogrupy R1a (słowiańskie i
indoirańskie). Śladem tego, że odmiana kentumowa była
wcześniejszą przed satemową formą języka
praindoeuropejskiego jest np. kentumowa postać starożytnego
języka tocharskiego w Zachodnich Chinach. Badania genetyczne Li et al. 2010 archeologicznych
szczątków siedmiu mężczyzn w Xiaohe (Xinjiang),
identyfikowanych, choć niepewnie, z Tocharami, wykazały
obecność tylko haplogrupy R1a1a.
Do podziału
języka praindoeuropejskiego doszło więc dopiero wtedy, gdy
synowskie haplogrupy R1a i R1b rozeszły się demograficznie i
terytorialnie. Ten podział populacji musiał nastąpić wnet
po powstaniu haplogrupy R1b, około 16000 lat temu, skoro nie
doszło do ich znacznego zmieszania się co do Y-DNA.
Określenie więc "Indoeuropejczycy", a raczej bardziej
logicznie: "Indo-Europejczycy", wskazujące dotąd tylko na
geograficzną rozległość pokrewnych języków i ich
użytkowników, jest teraz uzasadnione także pierwotną jednością
ich języka i wspólnym pochodzeniem biologicznym jego twórców.
Ojczyzny zaistnienia i
początkowego rozwoju wspólnoty praindoeuropejskiej trzeba szukać
zwłaszcza tam, gdzie pozostaje do dziś największe
zagęszczenie ojcowskie haplogrupy R1-M173. Może to być np. w
Azji Środkowej, począwszy od północnowschodniego Iranu, poprzez
pasmo Ataju i pogranicze Południowej Syberii, Mongolii i Chin. Np. obecne
zagęszczenie haplogrupy R1*, sięgające do 20% (Derenko et al. 2006) w niektórych
starych plemionach, np. Teleuci, Szorowie i Tofalarowie w Południowej
Syberii pod Ałtajem (u źródeł rzeki Ob i Jenisej) pod
granicą z Chinami i Mongolią świadczyłoby raczej za tym
regionem, choć później językowo dość gruntownie
sturkizowanym (zob. też Dyskusja n.10)
|
|
Drzewo genealogiczne głównych języków
indoeuropejskich |
A skąd zaś pochodzą języki
haplogrupy R1 czyli indoeuropejskie?
|
kliknij obraz, |
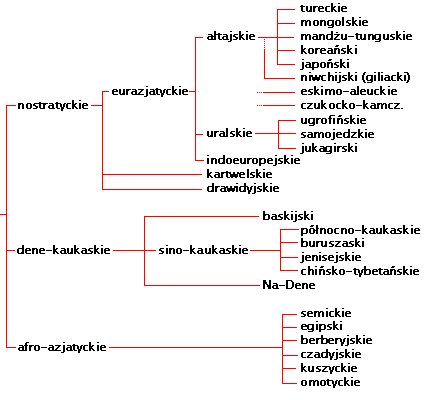
Języki
nostratyckie i ich drzewo genealogiczne
Oczywiści języki satem i kentum, i w ogóle praindoeuropejski
nie powstały nagle, lecz na drodze długiej ewolucji w
populacjach przodków. Cofając się dalej drogą ewolucji języków
w przeszłość, dochodzimy do rodziny języków
nostratyckich, które są językami ojcowskiej haplogrupy
NOP, przed jej rozgałęzieniem się i wyłonieniem z
haplogrupy P bliższej nam haplogrupy R, a potem z niej - haplogrupy R1
(indoeuropejskiej) i jej podgrup R1a (satem) i R1b (kentum). |
{kind=link}
Przeciwko korelacji starożytnych haplogrup chromosomowych Y-DNA z
językami nie może świadczyć fakt niektórych niekonsekwencji
w tej dziedzinie. Np. Baskowie, wyróżniający się haplogrupą
R1b1b2, zatraciły swój pierwotny język indoeuropejski, co mogło
się stać np. na skutek wyjątkowej inicjatywy strony
żeńskiej w pierwotnej małej grupie rodzinnej. Podobnie -
żyjąca w Basenie Dunaju ludność, wyróżniająca
się haplogrupami R1a1a, I2a, R1b1b2 i E (V-13), przyjęła
język małej grupy wojennej Madziarów (czyli przybyłych ze
wschodu Ugrów=Węgrów) z haplogrupą N; a mogło do tego
dojść na skutek prężnej polityki kulturowej zdobywców tej
ziemi wobec braku jednolitości językowej podbitej ludności
wieloplemiennej. Podobnie można wytłumaczyć kształt
greckiego języka indoeuropejskiego, użytkowanego głównie przez
przybyłe do środowiska europejskiego na Bałkanach populacje
haplogrupy E1b1b1 o genezie afrykańskiej; ciekawa jest wersja tego
języka, bo bliska indoirańskiemu, ale bez satemizacji. Podobnie
język Albańczyków, wyróżniających się głównie
haplogrupą R1b1b2, jest satemowy, co jest zapewne spowodowane bardziej
pierwotnym osadnictwem słowiańskim w tym rejonie i aktualnym
otoczeniem.
Interesujące
jest, że język siostrzanej względem R1 haplogrupy R2 w
południowych Indiach, drawidyjski, jest uważany za pokrewny
językom indoeuropejskim w ramach tzw. grupy języków nostratyckich
(zob. obraz wyżej). Bardziej dogłębne badania mogłyby
prajęzyków ludności haplogrupy R2 (w Azji Centralnej i w Indiach)
mogłyby okazać sie pożyteczne w tej kwestii.
|
Zasadniczo nie są znane przyczyny rozejścia się dróg
migracyjnych krewniaczych haplogrup R1a i R1b, a właściwie ich
podgrup, oznaczanych głównie jako R1a1 i R1b1b2, z Azji Centralnej ku
Europie. Genetyka, jak już wiemy (zob. wyżej, pkt 1),
częściowo zna mechanizmy powstawania mutacji, spontanicznych, a
także spowodowanych przyczynami zewnętrznymi, czyli mutagenami,
często niebezpiecznymi dla zdrowia, a nawet istnienia całej
potomnej populacji. |
W każdym razie wraz z oddaleniem się od siebie tych dwóch populacji,
dryfowały także i ich języki, różnicując się
wolniej wśród grupy wschodnioeuropejskiej R1a, a bardziej intensywnie
wśród zachodnioeuropejskiej grupy R1b, co było uzależnione
zwłaszcza od bardzo zróżnicowanych warunków życia. Duży
wpływ na późniejszy kształt języków południowych,
zachodnich i północnych w Europie miały bliskie kontakty i mieszanie
się ludności słowiańskiej R1a1 i celtyckiej R1b1b2 z
wcześniejszą, tzw. staroeuropejską, czyli z populacjami
weneckimi haplogrupy I (I1 i I2). W południowej Skandynawii doprowadziło
to do kreolizacji języków i powstania języka pragermańskiego.
Zapewne też znaczny był wpływ tzw. Staroeuropejczyków na
pozostałe języki grupy celtyckiej czy italskiej. A jaki wpływ
mieli nosiciele dynarskiej haplogrupy I2a2 na kształt języków słowiańskich? Podobnie
można by śledzić wpływ języków nieindoeuropejskich
populacji bliskowschodnich J1 i J2, kaukaskiej G2a i
północno-wschodnioafrykańskiej E, które do Europy, zwłaszcza
południowej, napływały w mezolicie lub na początku neolitu
(zob. niżej).
Także w populacji indyjskich Ariów, liczącej
dziś około 100 milionów ludzi z haplotypem słowiańskim
R1a1a (16% społeczeństwa Indii), powstały nowe, daleko
idące modyfikacje językowe
prasłowiańsko-praindoirańskie. W tym języku, zwanym
sanskrytem wedyjskim, ułożono przed trzema tysiącami lat w
Pendżabie święte hymny
Rygwedy, wzywające bogów do udzielenia pomocy
wojowniczym Ariom w walce z lokalnymi wrogami (zapewne plemionami
drawidyjskimi). W tych tekstach, wiernie przekazywanych ustnie od
tysiącleci do czasu ich spisania około XIV w. po Chr., wyczuwa
się reminiscencję dokonanego przez Ariów podboju. Populacja R1a1a w
Indiach była najprawdopodobniej twórcą także systemu kastowego;
w najwyższej kaście, wśród braminów, występują oni do
około 72%!
Postscriptum o teorii Dawida
Anthony w książce: Koń,
koło i język, zob. Dyskusja n.12
Zob. także o problemie,
która haplogrupa jest cechą Indoeuropejczyków Dyskusja n.14.
(Nienaukowa więc, krzywdząca dla innych ludów europejskich, a
zarazem bardzo szkodliwa dla szacunku Polaków w Europie, jest teza, szerzona na
niektórych portalach www. o tożsamości Indoeuropejczyków tylko ze
Słowianami, wywyższanie języka i kultury Słowian, ich
siły i rzekomej wojowniczości, z wyraźną nutką
szowinizmu i pogardy dla innych narodów, które rzekomo tylko dzięki
Słowianom stały się indoeuropejskie. Podobne głosy odzywają
sie i w niektórych środowiskach nacjonalistów Rosji czy Ukrainy. Jest to
próba zastąpienia głupiego przedwojennego pangermanizmu niemieckiego
równie głupim i niebezpiecznym panslawizmem genetycznym).